

 夜黎
夜黎本文章內容基於萊斯大學 OpenStax 的 Anatomy and Physiology 2e,由夜黎重新編輯。(根據本書前言中的創用 CC BY 4.0 聲明)
原文傳送門:<3> The Cellular Level of Organization — 3.1 The Cell Membrane
索引傳送門:《解剖學和生理學2e》索引頁面
學習本章後,你將能夠:
- 描述構成細胞膜的分子成分
- 解釋細胞膜的主要特徵和特性
- 區分可以和不能通過脂質雙層〔lipid bilayer〕擴散的材料
- 比較和對比不同類型的被動傳輸與主動傳輸,並提供每種類型的例子
前言
儘管結構和功能存在差異,多細胞生物中的所有活細胞都具有周圍的細胞膜。 由於皮膚的外層將你的身體與其環境分開,細胞膜〔cell membrane〕(也稱為質膜〔plasma membrane〕)將細胞的內部內容與其外部環境分開。 這種細胞膜在細胞周圍提供了保護屏障,並調節哪些物質可以進出。
細胞膜的結構和組成
細胞膜〔cell membrane〕是一種極其柔韌的結構,主要由背對背磷脂〔back-to-back phospholipids〕(“雙層〔bilayer〕”)組成。 還存在膽固醇,它有助於膜的流動性,並且膜內嵌入有各種具有多種功能的蛋白質。
單個磷脂分子的一端有一個磷酸基團,稱為“頭〔head〕”,兩條並排的脂肪酸鏈構成了脂質尾(圖 3.2)。 磷酸基團帶負電,使頭部具有極性和親水性,或“愛水〔water loving〕”。 親水分子〔hydrophilic molecule〕(或分子區域)是被水吸引的分子。 因此,磷酸鹽頭〔phosphate heads〕被細胞外和細胞內環境的水分子吸引。 另一方面,脂質尾部不帶電荷,或非極性,並且是疏水性的,或“怕水〔water fearing〕”。 疏水性分子〔hydrophobic molecule〕 (或分子區域)排斥水並被水排斥。 有些脂尾由飽和脂肪酸組成,有些則含有不飽和脂肪酸。 這種組合增加了不斷運動的尾巴的流動性。 因此磷脂是兩親性分子。 兩親性分子〔amphipathic molecule〕是同時包含親水性和疏水性區域的分子。 事實上,肥皂可以去除油污和油漬,因為它具有兩親性。 親水部分可以溶解在水中,而疏水部分可以將油脂捕獲在膠束中,然後可以將其洗掉。
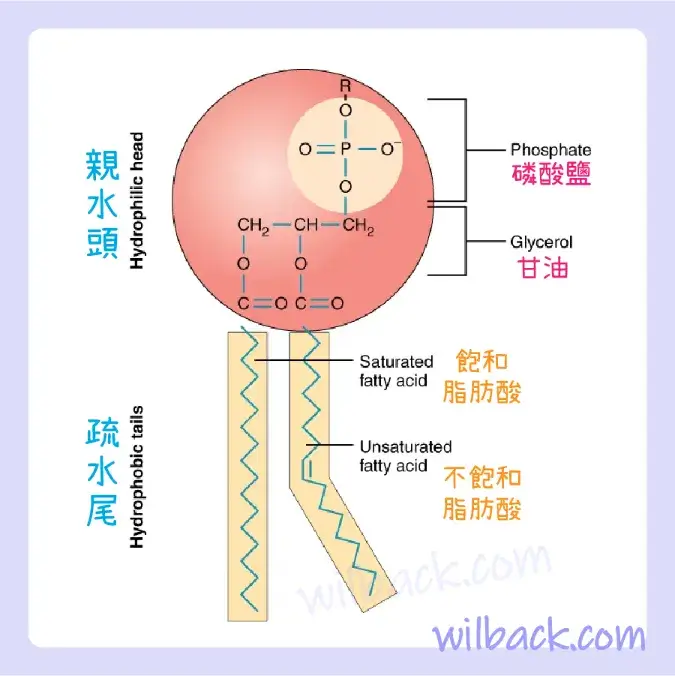
磷脂分子由親水性的極性磷酸鹽“頭”和疏水性的非極性脂質“尾”組成。 不飽和脂肪酸會導致疏水尾部扭結。
細胞膜由兩層相鄰的磷脂層組成。 一層的脂質尾部面向另一層的脂質尾部,在兩層的界面處相遇。 磷脂頭朝外,一層暴露在細胞內部,一層暴露在外部(圖3.3)。 由於磷酸基團具有極性和親水性,因此它們會被細胞內液中的水吸引。 細胞內液〔intracellular fluid〕(ICF)是細胞內部的液體。 磷酸基團也被細胞外液吸引。 細胞外液〔extracellular fluid〕(ECF)是細胞膜外殼外部的液體環境。 間質液〔interstitial fluid〕 (IF) 是指不包含在血管內的細胞外液。 由於脂質尾部是疏水性的,它們在膜的內部區域相遇,從該空間排除含水的細胞內液和細胞外液。 細胞膜含有許多與磷脂雙層相關的蛋白質以及其他脂質(例如膽固醇)。 膜的一個重要特點是它保持流動性;細胞膜中的脂質和蛋白質並不僵硬地固定在一個位置。
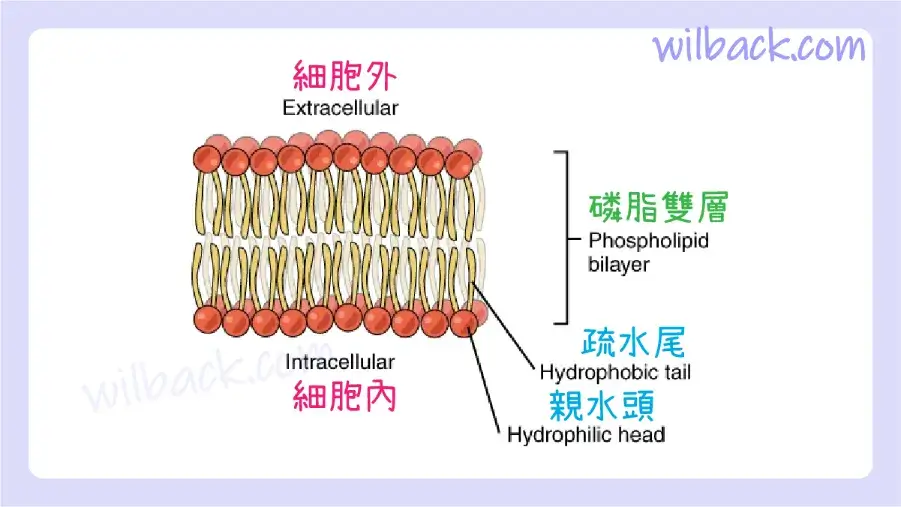
磷脂雙層由兩個相鄰的磷脂片組成,尾對尾排列。 疏水尾部彼此相連,形成膜的內部。 極性頭接觸細胞內部和外部的液體。
膜蛋白
脂質雙層構成細胞膜的基礎,但其內部佈滿了各種蛋白質。 通常與細胞膜相關的兩種不同類型的蛋白質是嵌入蛋白〔integral protein〕和周邊蛋白〔peripheral protein〕(圖 3.4)。 顧名思義,嵌入蛋白是嵌入膜中的蛋白質。 通道蛋白〔channel protein〕是嵌入蛋白的一個例子,它選擇性地允許特定物質(例如某些離子)進出細胞。
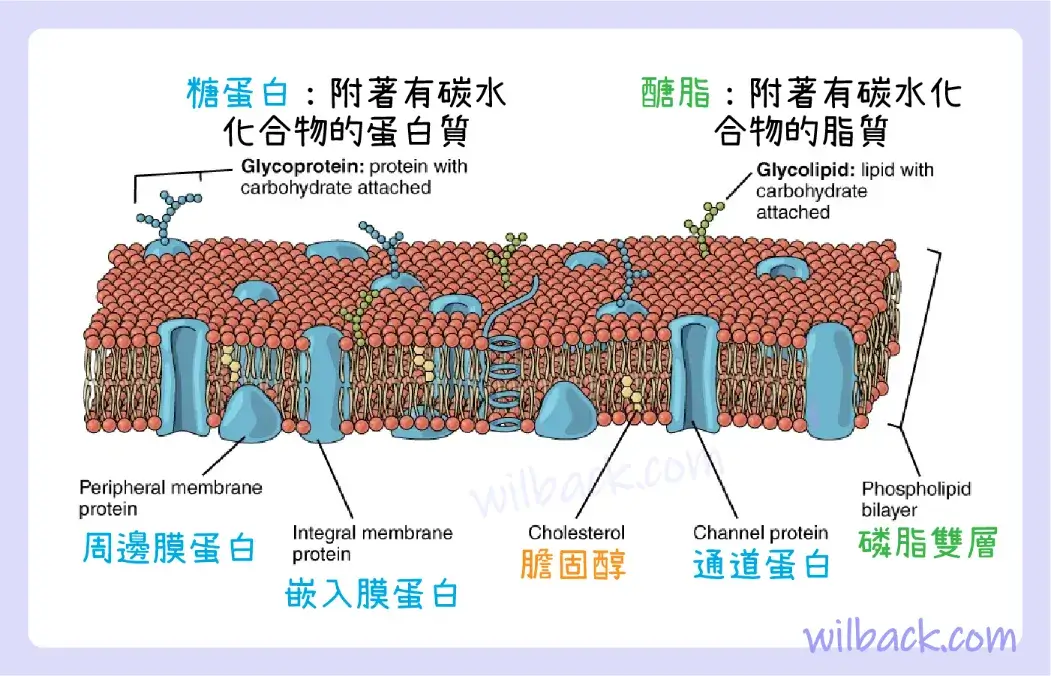
細胞的細胞膜是磷脂雙層,含有許多不同的分子成分,包括蛋白質和膽固醇,其中一些還附有碳水化合物基團。
另一組重要的整合蛋白是細胞識別蛋白〔cell recognition proteins〕,它用於標記細胞的身份,以便它可以被其他細胞識別。 受體〔receptor〕是一類識別蛋白,可以選擇性地結合細胞外的特定分子,這種結合誘導細胞內的化學反應。 配體〔ligand〕是結合並且激活受體的特定分子。 一些嵌入蛋白具有受體和離子通道的雙重作用。 受體-配體相互作用的一個例子是神經細胞上結合神經遞質(例如多巴胺〔dopamine〕)的受體。 當多巴胺分子與多巴胺受體蛋白結合時,跨膜蛋白〔transmembrane protein〕內的通道打開,允許某些離子流入細胞。
一些嵌入膜蛋白是糖蛋白。 糖蛋白〔glycoproteins〕是一種附著有碳水化合物分子的蛋白質,其延伸到細胞外基質〔extracellular matrix〕中。 糖蛋白上附著的碳水化合物標籤有助於細胞識別。 從膜蛋白延伸出來,甚至從一些膜脂質〔membrane lipids〕延伸出來的碳水化合物共同形成糖萼。 糖萼〔glycocalyx〕是細胞周圍的一層看起來模糊的塗層,由附著在細胞膜上的糖蛋白和其他碳水化合物形成。 糖萼可以具有多種作用。 例如,它可能含有允許細胞與另一個細胞結合的分子,它可能含有激素受體,或者可能含有分解營養物質的酶。 人體內發現的糖萼是該人基因構成的產物。 它們賦予每個人數万億個細胞歸屬於人體的“身份〔identity〕”。 這種身份是一個人的免疫防禦細胞“知道”不攻擊自己的身體細胞的主要方式,但這也是另一個人捐贈的器官可能被拒絕的原因。
周邊蛋白〔peripheral proteins〕通常存在於脂雙層的內表面或外表面上,但也可以附著於嵌入蛋白的內表面或外表面。 這些蛋白質通常對細胞執行特定的功能。 例如,腸細胞表面的一些周邊蛋白質充當消化酶,將營養物質分解成可以通過細胞並進入血液的尺寸。
跨細胞膜運輸
細胞膜的偉大奇蹟之一是它能夠調節細胞內物質的濃度。 這些物質包括Ca++、Na+、K+和Cl–等離子; 營養素,包括糖、脂肪酸和氨基酸; 以及必須離開細胞的廢物,特別是二氧化碳 (CO2)。
膜的脂質雙層結構提供了第一級控制。 磷脂緊密地堆積在一起,膜具有疏水性內部。 這種結構使膜具有選擇性滲透性。 具有選擇性滲透性〔selective permeability〕的膜只允許滿足特定標準的物質在沒有幫助的情況下通過它。 就細胞膜而言,只有相對較小的非極性材料可以穿過脂質雙層(請記住,膜的脂質尾部是非極性的)。 其中的一些例子是其他脂質、氧氣和二氧化碳氣體以及酒精。 然而,水溶性材料(如葡萄糖、氨基酸和電解質)需要一些幫助才能穿過膜,因為它們會被磷脂雙層的疏水尾部排斥。 所有穿過膜的物質都是通過兩種通用方法之一進行的,這兩種方法根據是否需要能量進行分類。 被動運輸〔passive transport〕是指物質在不消耗細胞能量的情況下穿過細胞膜的運動。 相比之下,主動運輸〔active transport〕是物質利用三磷酸腺苷 (ATP) 的能量跨膜移動。
被動運輸
為了了解物質如何被動地穿過細胞膜,有必要了解濃度梯度和擴散。 濃度梯度〔concentration gradient〕是物質在空間中的濃度差異。 分子(或離子)將從它們較集中的地方擴散/擴散〔spread/diffuse〕到它們較不集中的地方,直到它們均勻地分佈在該空間中。 (當分子以這種方式移動時,據說它們會沿著濃度梯度移動。) 擴散〔diffusion〕是指粒子從濃度較高的區域移動到濃度較低的區域。 幾個常見的例子將有助於說明這個概念。 想像一下在一個封閉的浴室內。 如果噴一瓶香水,香味分子會自然地從離開瓶子的地方擴散到浴室的各個角落,這種擴散會一直持續到不再有濃度梯度為止。 另一個例子是將一勺糖放入一杯茶中。 最終,糖會擴散到整個茶中,直到不存在濃度梯度。 在這兩種情況下,如果房間較暖或茶較熱,擴散會發生得更快,因為分子相互碰撞並擴散得比在較冷的溫度下更快。 因此,體內溫度約為 98.6°F 也有助於體內顆粒的擴散。
每當一種物質以較高濃度存在於半透膜〔semipermeable membrane〕(例如細胞膜)的一側時,可以沿膜濃度梯度向下移動的任何物質都會這樣做。 考慮可以輕鬆穿過細胞膜脂質雙層的物質,例如氧氣 (O2) 和 CO2。 O2 通常會擴散到細胞中,因為它在細胞外部更集中,而 CO2 通常會擴散到細胞外,因為它在細胞內部更集中。 這些例子都不需要細胞提供任何能量,因此它們使用被動運輸跨膜移動。
在繼續之前,你需要檢查可以擴散穿過細胞膜的氣體。 由於細胞在新陳代謝過程中會迅速消耗氧氣,因此細胞內部的氧氣濃度通常低於外部。 結果,氧氣將從間質液〔interstitial fluid〕中直接擴散通過膜的脂質雙層並進入細胞內的細胞質。 另一方面,由於細胞產生CO2作為新陳代謝的副產品,因此細胞質內的CO2濃度升高; 因此,CO2將從細胞穿過脂質雙層進入組織液,在組織液中其濃度較低。 這種分子穿過細胞膜從濃度較高的一側移動到濃度較低的一側的機制是一種被動運輸形式,稱為簡單擴散〔simple diffusion〕(圖 3.5)。
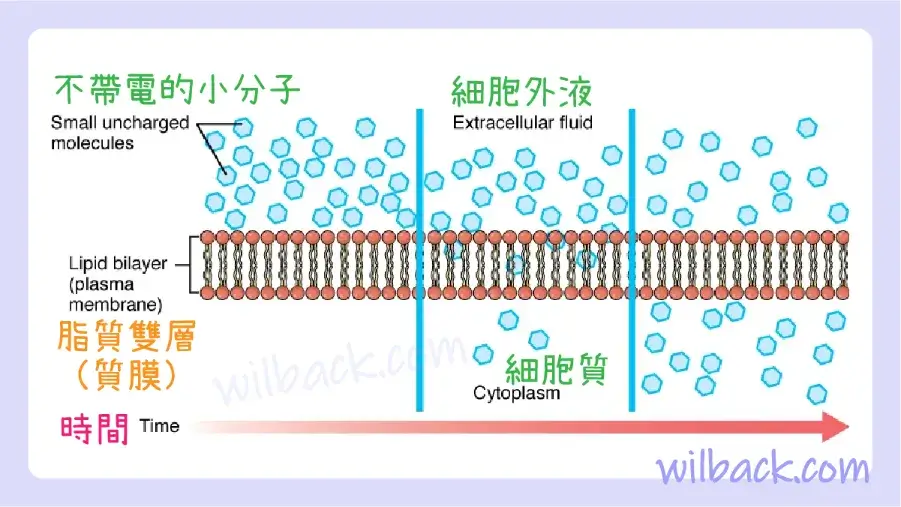
脂質雙層的結構允許小的、不帶電的物質(例如氧氣和二氧化碳)以及疏水性分子(例如脂質)通過簡單擴散沿著其濃度梯度穿過細胞膜。
親水性的大極性或離子分子不能輕易穿過磷脂雙層。 非常小的極性分子,例如水,由於尺寸小,可以通過簡單的擴散來穿過。 任何大小的帶電原子或分子都不能通過簡單的擴散穿過細胞膜,因為電荷被磷脂雙層內部的疏水尾部排斥。 溶解在細胞膜兩側水中的溶質往往會沿著其濃度梯度擴散,但由於大多數物質不能自由通過細胞膜的脂質雙層,因此它們的運動僅限於蛋白質通道和膜中的專門運輸機制。 促進擴散是用於那些由於其尺寸、電荷和/或極性而無法穿過脂質雙層的物質的擴散過程(圖 3.6)。 促進擴散〔Facilitated diffusion〕的一個常見例子是葡萄糖進入細胞,並在細胞中用於製造 ATP。 儘管葡萄糖在細胞外可以更加濃縮,但它不能通過簡單擴散穿過脂質雙層,因為它既大又具極性。 為了解決這個問題,一種稱為葡萄糖轉運蛋白〔glucose transporter〕的特殊載體蛋白會將葡萄糖分子轉移到細胞中,以促進其向內擴散。
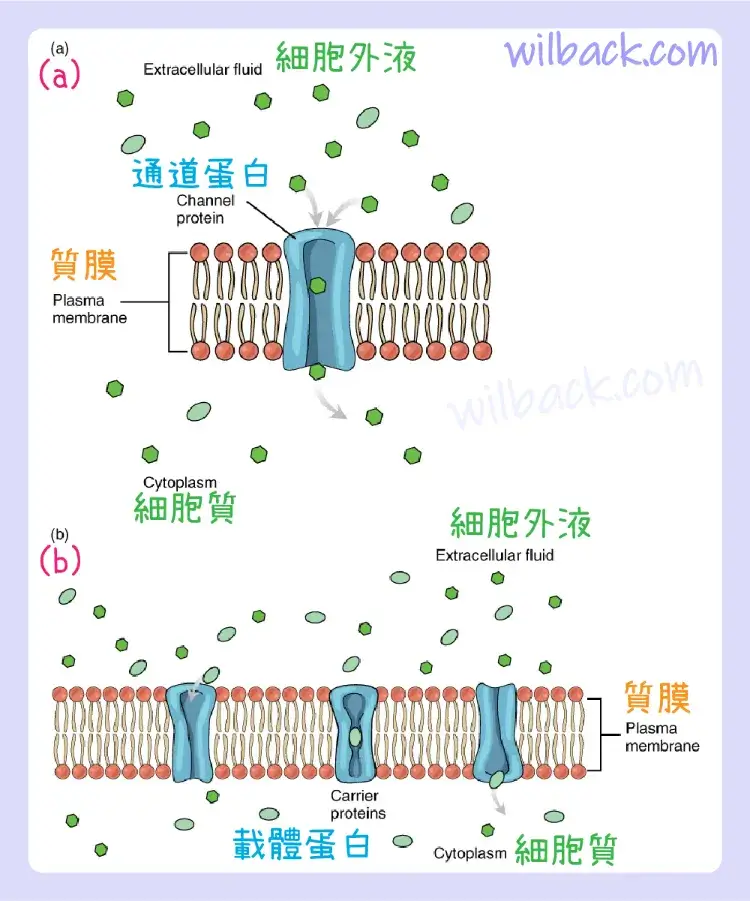
(a) 物質穿過細胞(質)膜的促進擴散是在通道蛋白和載體蛋白等蛋白質的幫助下發生的。 通道蛋白的選擇性低於載體蛋白,並且通常根據大小和電荷稍微區分其貨物。 (b) 載體蛋白更具選擇性,通常只允許一種特定類型的分子交叉。
例如,即使鈉離子 (Na+) 在細胞外高度濃縮,這些電解質也會帶電並且無法穿過細胞膜的非極性脂質雙層。 形成鈉通道(或“孔〔pores〕”)的膜蛋白促進它們的擴散,因此 Na+ 離子可以沿著濃度梯度從細胞外移動到細胞內。 還有許多其他溶質必須經過促進擴散才能進入細胞,例如氨基酸,或離開細胞,例如廢物。 由於促進擴散是一個被動過程,因此不需要細胞消耗能量。
水還可以通過蛋白質通道或通過膜本身的脂質尾部之間的滑動,自由地穿過所有細胞的細胞膜。 滲透〔osmosis〕是水通過半透膜〔semipermeable membrane〕的擴散(圖 3.7)。
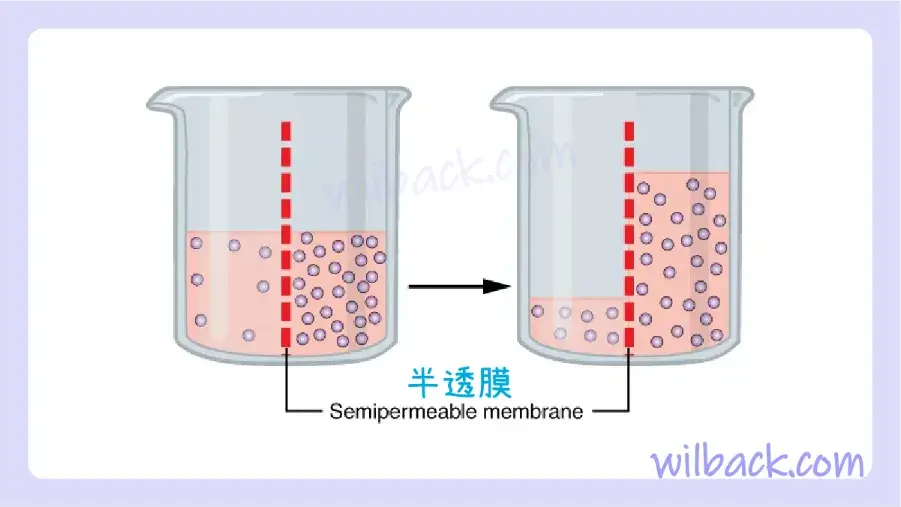
滲透是水通過半透膜沿著濃度梯度擴散。 如果膜可滲透水,但不可滲透溶質,則水將通過擴散到水濃度較低的一側(以及因此溶質濃度較高的一側)來平衡其自身濃度。 在左邊的燒杯中,膜右側的溶液是高滲的。
水分子的運動本身不受某些細胞的調節,因此重要的是這些細胞暴露在一個外部溶質濃度(在細胞外液中)等於細胞內溶質濃度(在細胞質中)的環境中。 具有相同溶質濃度的兩種溶液被稱為等張〔isotonic〕(等張力〔equal tension〕)。 當細胞與其細胞外環境等滲時,細胞內外水分子的濃度相同,細胞保持正常的形狀(和功能)。
當細胞外部與細胞內部的溶質不平衡時,就會發生滲透〔osmosis〕。 溶質濃度高於另一種溶液的溶液被稱為高張溶液〔hypertonic solution〕,水分子傾向於擴散到高張溶液中(圖 3.8)。 當水通過滲透作用離開細胞時,高滲透溶液中的細胞會萎縮。 相反,溶質濃度低於另一種溶液的溶液被稱為低張溶液〔hypotonic solution〕,水分子傾向於從低張溶液中擴散出來。 低張溶液中的細胞會吸收過多的水並膨脹,最終有破裂的風險。 生物體內平衡的一個關鍵方面是創造一個內部環境,使身體的所有細胞都處於等滲溶液中。 各種器官系統,特別是腎臟,致力於維持這種體內平衡。
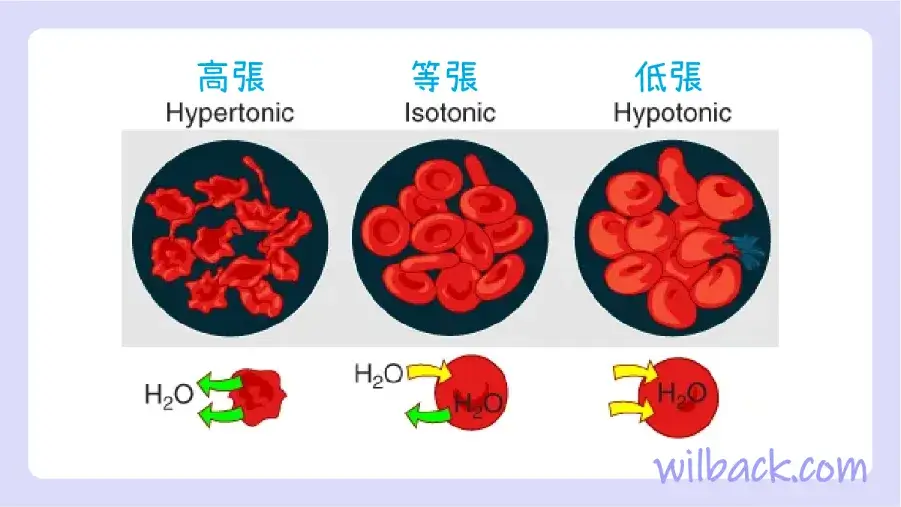
高張溶液的溶質濃度高於另一種溶液。 等張溶液的溶質濃度等於另一種溶液。 低張溶液的溶質濃度低於另一種溶液。
除了擴散之外,在隔室之間被動運輸材料的另一種機制是過濾〔filtration〕。 與物質從濃度較高的地方擴散到濃度較低的地方不同,過濾使用靜水壓力梯度〔hydrostatic pressure gradient〕,將流體及其中的溶質從高壓區域推向低壓區域。 過濾是體內極其重要的過程。 例如,循環系統利用過濾將血漿和物質穿過毛細血管內皮層並進入周圍組織,為細胞提供營養。 腎臟的過濾壓力提供了從血液中清除廢物的機制。
主動運輸
對於上述所有運輸方法,細胞不消耗能量。 幫助物質被動運輸的膜蛋白無需使用 ATP 即可實現這一目的。 在主動運輸過程中,ATP 需要將物質穿過膜,通常需要蛋白質載體的幫助,並且通常逆其濃度梯度。
最常見的主動運輸類型之一涉及充當幫浦的蛋白質。 “幫浦〔pump〕”這個詞可能會讓人想到利用能量給自行車或籃球的輪胎打氣。 同樣,這些膜蛋白需要來自 ATP 的能量來跨膜運輸物質(分子或離子),通常是逆著濃度梯度(從低濃度區域到高濃度區域)。
鈉鉀幫浦〔sodium-potassium pump〕,也稱為 Na+/K+ ATP 酶〔Na+/K+-ATPase〕,將鈉輸送出細胞,同時將鉀移入細胞。 Na+/K+ 幫浦是多種細胞膜中發現的重要離子幫浦。 這些幫浦在神經細胞中特別豐富,神經細胞不斷幫浦出鈉離子並吸入鉀離子,以維持細胞膜上的電梯度。 電梯度〔electrical gradient〕是空間中電荷的差異。 例如,就神經細胞而言,細胞內部和外部之間存在電梯度,內部相對於外部帶負電(約 -70 mV)。 負電梯度得以維持,因為每個 Na+/K+ 幫浦將三個 Na+ 離子移出細胞,並將兩個 K+ 離子移入每個使用的 ATP 分子的細胞中(圖 3.9)。 這個過程對於神經細胞來說非常重要,它佔據了神經細胞 ATP 使用的大部分。
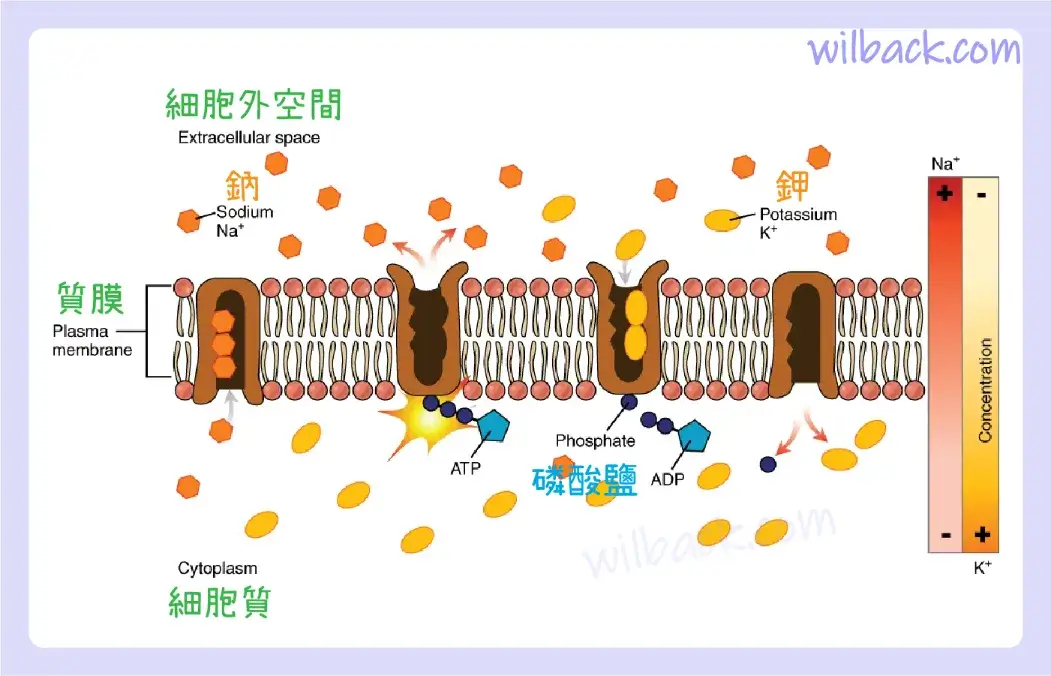
鈉鉀幫浦存在於許多細胞(質)膜中。 在 ATP 的驅動下,幫浦將鈉離子和鉀離子向相反的方向移動,每個方向都逆著其濃度梯度。 在幫浦的單個循環中,三個鈉離子被擠出細胞,兩個鉀離子被輸入到細胞中。
主動運輸幫浦還可以與其他主動或被動運輸系統一起工作,將物質跨膜移動。 例如,鈉鉀幫浦在細胞外維持高濃度的鈉離子。 因此,如果細胞需要鈉離子,它所要做的就是打開一個被動的鈉通道,因為鈉離子的濃度梯度會驅使它們擴散到細胞中。 通過這種方式,主動傳輸幫浦(鈉鉀幫浦)的作用通過產生濃度梯度為鈉離子的被動傳輸提供動力。 當主動運輸以這種方式為另一種物質的運輸提供動力時,稱為二次主動運輸〔secondary active transport〕。
同向轉運者〔symporters〕是沿同一方向移動兩種物質的次級主動轉運者。 例如,鈉-葡萄糖同向轉運者使用鈉離子將葡萄糖分子“拉”入細胞。 由於細胞儲存葡萄糖作為能量,因此細胞內部的葡萄糖濃度通常高於細胞外部的濃度。 然而,由於鈉鉀幫浦的作用,當同向轉運蛋白打開時,鈉離子很容易擴散到細胞內。 通過同向轉運者的大量鈉離子提供了能量,使葡萄糖逆其濃度梯度穿過同向轉運者並進入細胞。
相反,反向轉運者〔antiporters〕是沿相反方向轉運物質的次級主動轉運系統。 例如,鈉-氫離子反向轉運者利用鈉離子向內湧入的能量將氫離子 (H+) 移出細胞。 鈉氫逆向轉運者用於維持細胞內部的 pH 值。
其他形式的主動運輸不涉及膜載體。 內吞作用〔endocytosis〕(“進入細胞”)是細胞通過將物質包裹在細胞膜的一部分中,然後夾斷該部分膜來攝取物質的過程(圖 3.10)。 一旦被夾斷,膜的部分及其內容物就變成獨立的細胞內囊泡〔intracellular vesicle〕。 囊泡〔vesicle〕是一種膜囊〔membranous sac〕——一種由脂質雙層膜包圍的球形中空細胞器。 內吞作用通常會將必須分解或消化的物質帶入細胞。 吞噬作用〔phagocytosis〕(“細胞吞噬”)是大顆粒的內吞作用。 許多免疫細胞參與吞噬入侵病原體。 就像小吃豆人一樣,它們的工作是巡邏身體組織,尋找不需要的物質,例如入侵的細菌細胞,吞噬它們並消化它們。 與吞噬作用相反,胞飲作用 〔pinocytosis〕(“細胞飲用”)將含有溶解物質的液體通過膜囊泡帶入細胞。
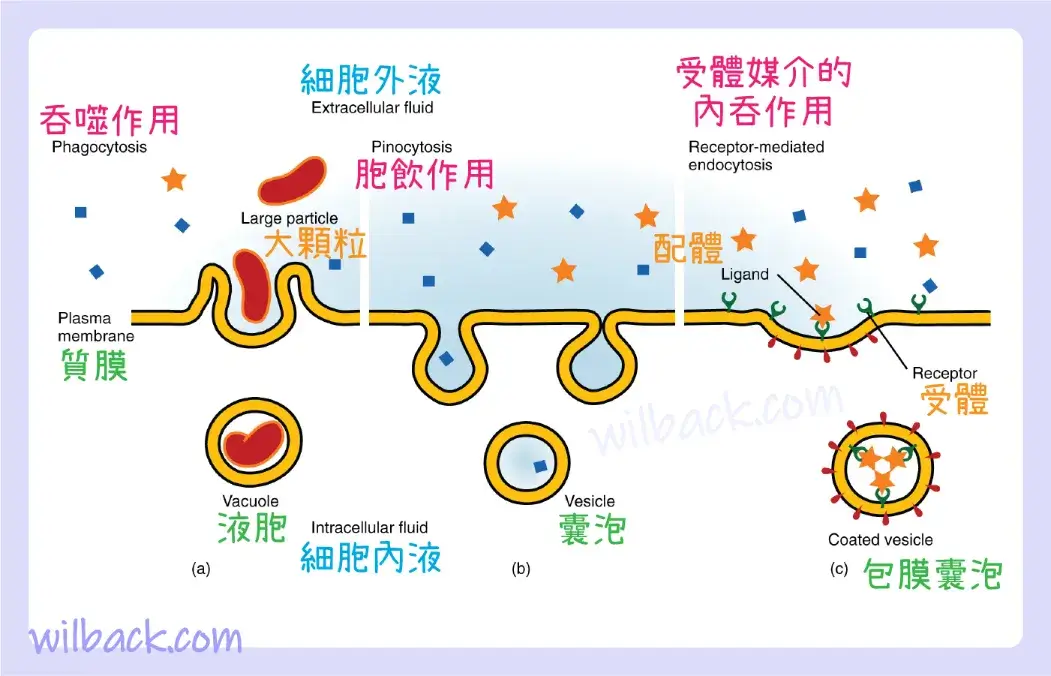
內吞作用是主動運輸的一種形式,其中細胞利用其細胞膜包裹細胞外物質。 (a) 在相對非選擇性的吞噬作用中,細胞攝入大顆粒。 (b) 在胞飲作用中,細胞吸收液體中的小顆粒。 (c) 相反,受體媒介的內吞作用具有相當的選擇性。 當外部受體結合特定配體時,細胞通過內吞該配體做出反應。
吞噬作用和胞飲作用吸收大部分細胞外物質,並且它們通常對引入的物質沒有高度選擇性。細胞通過受體介導的內吞作用來調節特定物質的內吞作用。 受體媒介的內吞作用〔receptor-mediated endocytosis〕是由細胞膜的一部分進行的內吞作用,該部分含有許多對某些物質具有特異性的受體。 一旦表面受體結合了足夠量的特定物質(受體的配體〔receptor’s ligand〕),細胞就會內吞含有受體-配體複合物的細胞膜部分。 鐵是血紅蛋白的必需成分,通過這種方式被紅血球內吞。 鐵與血液中一種稱為轉鐵蛋白〔transferrin〕的蛋白質結合。 紅血球表面上的特定轉鐵蛋白受體與鐵的轉鐵蛋白分子結合,細胞內吞受體-配體複合物。
與內吞作用相反,胞吐作用〔exocytosis〕(“從細胞中取出”)是細胞利用囊泡運輸輸出物質的過程(圖 3.11)。 許多細胞製造必須分泌的物質,就像製造出口產品的工廠一樣。 這些物質通常被包裝到細胞內的膜結合囊泡〔membrane-bound vesicles〕 中。 當囊泡膜與細胞膜融合時,囊泡將其內容物釋放到間質液中。 然後囊泡膜成為細胞膜的一部分。 胃和胰腺細胞通過胞吐作用產生和分泌消化酶(圖3.12)。 內分泌細胞產生並分泌輸送到全身的激素,某些免疫細胞產生並分泌大量組織胺〔histamine〕,這是一種對免疫反應很重要的化學物質。
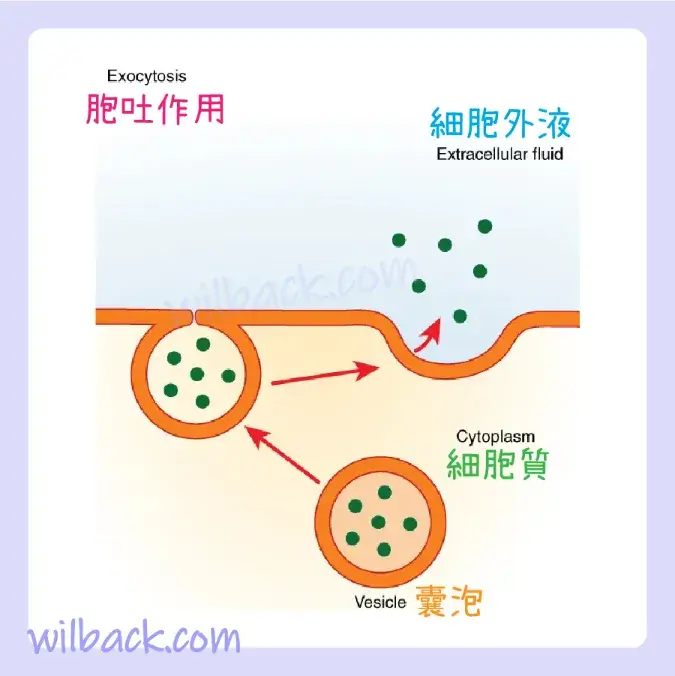
胞吐作用與內吞作用非常相似。 用於出口的材料被包裝到細胞內的囊泡中。 囊泡膜與細胞膜融合,內容物釋放到細胞外空間。
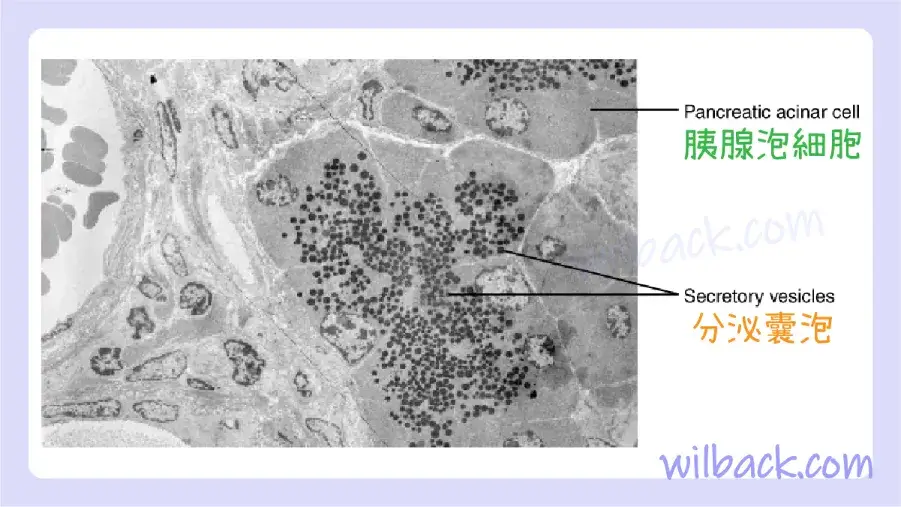
胰腺腺泡細胞產生並分泌許多消化食物的酶。 該電子顯微照片中的微小黑色顆粒是充滿酶的分泌囊泡,這些酶將通過胞吐作用從細胞中排出。 LM × 2900。(顯微照片由密歇根大學醫學院董事會提供 © 2012)
細胞:囊性纖維化〔Cystic Fibrosis〕
在美國,囊性纖維化 (CF) 影響著大約 30,000 人,每年報告大約 1,000 例新病例。 這種遺傳病最著名的是它對肺部的損害,導致呼吸困難和慢性肺部感染,但它也會影響肝臟、胰腺和腸道。 僅在大約 50 年前,出生時患有 CF 的兒童的預後非常嚴峻——預期壽命很少超過 10 年。 如今,隨著醫療技術的進步,許多CF患者都活到了30多歲。
CF 的症狀是由稱為囊性纖維化跨膜電導調節因子〔cystic fibrosis transmembrane conductance regulator〕 (CFTR) 的膜離子通道故障引起的。 在健康人中,CFTR 蛋白是一種完整的膜蛋白,可將 Cl- 離子轉運出細胞。 在患有 CF 的人中,CFTR 基因發生突變,因此細胞會產生有缺陷的通道蛋白,該蛋白通常不會併入膜中,而是被細胞降解。
CFTR 需要 ATP 才能發揮作用,使其 Cl- 運輸成為一種主動運輸形式。 這一特性長期以來讓研究人員感到困惑,因為 Cl- 離子在被輸送出細胞時實際上是沿著濃度梯度流動的。 主動傳輸通常會逆著濃度梯度幫浦送離子,但 CFTR 是該規則的一個例外。
在正常肺組織中,Cl- 移動出細胞,在細胞外維持富含 Cl-、帶負電的環境。 這對於呼吸系統的上皮襯裡尤其重要。 呼吸道上皮細胞分泌粘液,用於捕獲灰塵、細菌和其他碎片。 纖毛(複數=纖毛)是某些細胞上發現的毛髮狀附屬物之一。 上皮細胞上的纖毛將粘液及其捕獲的顆粒沿氣道移動,遠離肺部並向外移動。 為了有效向上移動,粘液不能太粘稠; 相反,它必須具有稀薄的水狀稠度。 Cl– 的運輸和細胞外負電環境的維持將 Na+ 等正離子吸引到細胞外空間。 Cl– 和 Na+ 離子在細胞外空間中積累,形成富含溶質的粘液,其中水分子濃度較低。 結果,通過滲透作用,水從細胞和細胞外基質轉移到粘液中,將其“稀釋”。 這就是在正常的呼吸系統中,粘液保持充分稀釋以被排出呼吸系統的方式。
如果CFTR通道不存在,Cl-離子就不會以足夠的數量被輸送出細胞,從而阻止它們吸引正離子。 分泌的粘液中缺乏離子導致缺乏正常的水濃度梯度。 因此,不存在將水拉入粘液的滲透壓。 產生的粘液又厚又粘,纖毛上皮無法有效地將其從呼吸系統中清除。 肺部通道被粘液及其攜帶的碎片堵塞。 由於細菌細胞無法有效地從肺部帶走,因此更容易發生細菌感染。
更新紀錄
2023/09/14 發佈本文
評論