

 夜黎
夜黎本文章內容基於萊斯大學 OpenStax 的 Anatomy and Physiology 2e,由夜黎重新編輯。(根據本書前言中的創用 CC BY 4.0 聲明)
原文傳送門:<10> Muscle Tissue — 10.4 Nervous System Control of Muscle Tension
索引傳送門:《解剖學和生理學2e》索引頁面
學習本節後,你將能夠:
- 解釋向心、等長和離心收縮
- 描述長度與張力的關係
- 描述肌肉抽搐的三個階段
- 定義波加成〔wave summation〕、強直〔tetanus〕和階梯現象〔treppe〕
前言
為了移動物體(稱為負荷〔load〕),骨骼肌肌纖維中的肌節必須縮短。 肌肉收縮(或肌節縮短)所產生的力量稱為肌張力〔muscle tension〕。 然而,當肌肉抵抗不移動的負荷而收縮時,也會產生肌張力,導致骨骼肌收縮的兩種主要類型:等張收縮〔isotonic contractions〕和等長收縮〔isometric contractions〕。
在等張收縮〔isotonic contractions〕中,肌肉的張力保持恆定,負荷隨著肌肉長度的變化(縮短)而移動。 等張收縮有兩種:向心收縮和離心收縮。 同心收縮〔concentric contraction〕涉及肌肉縮短以移動負荷。 一個例子是,當手的重量隨著肌肉張力的增加而向上提升時,肱二頭肌會收縮。 當肱二頭肌收縮時,肘關節的角度會隨著前臂靠近身體而減少。 在這裡,肱二頭肌隨著肌纖維中的肌節縮短,並形成橫橋而收縮; 肌球蛋白頭〔myosin heads〕拉動肌動蛋白〔actin〕。 當肌肉張力減弱且肌肉拉長時,會發生離心收縮〔eccentric contraction〕。 在這種情況下,隨著神經系統刺激激活的橫橋數量減少,手的重量以緩慢且受控的方式降低。 在這種情況下,隨著肱二頭肌的張力釋放,肘關節的角度增加。 離心收縮也用於身體的運動和平衡。
當肌肉產生張力而不改變骨骼關節的角度時,會發生等長收縮〔isometric contraction〕。 等長收縮涉及肌節縮短和增加的肌肉張力,但不會移動負荷,因為產生的力無法克服負荷提供的阻力。 例如,如果一個人試圖舉起過重的手部重量,則會出現肌節激活並縮短至一定程度,肌肉張力不斷增加,但肘關節角度沒有變化。 在日常生活中,等長收縮在〝維持姿勢〞以及〝維持骨骼和關節的穩定性〞中非常活躍。 然而,將頭部保持直立並不是因為肌肉無法移動頭部,而是因為目標是保持靜止而不產生運動。 身體的大多數動作都是由於等張收縮和等長收縮共同作用,從而產生廣泛的結果(圖10.13)。
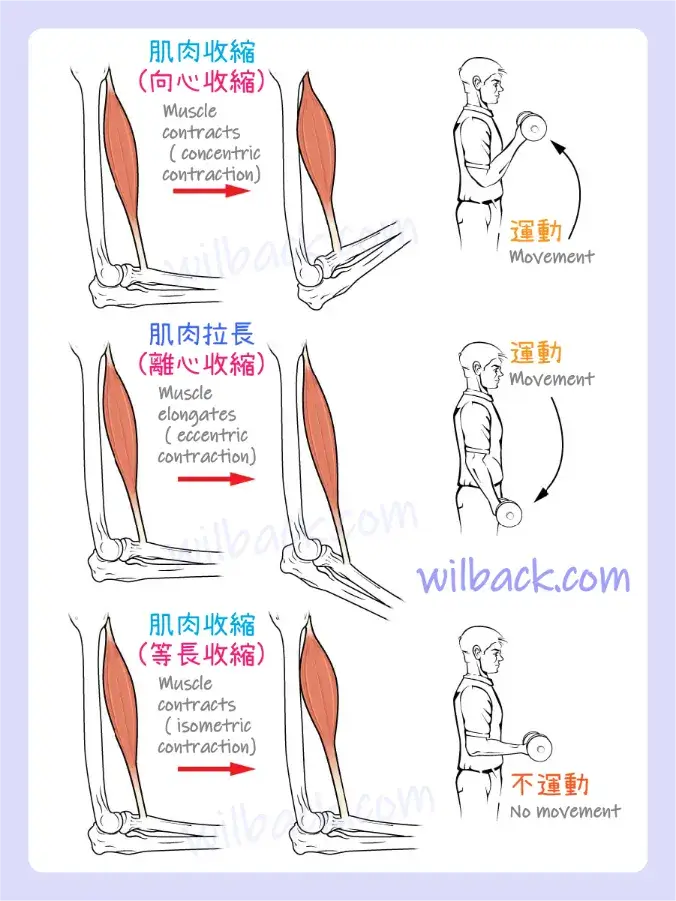
在等張收縮〔isotonic contractions〕期間,肌肉長度會改變以移動負荷。 在等長收縮〔isometric contractions〕期間,肌肉長度不會改變,因為負荷超過了肌肉所能產生的張力。
所有這些肌肉活動都在神經系統的精確控制之下。 神經控制調節向心、離心和等長收縮、肌纖維募集和肌張力。 神經系統控制骨骼肌的一個重要面向是〝運動單位的作用〞。
運動單位
如你所學到的,每根骨骼肌纖維都必須受到運動神經元軸突末端的支配才能收縮。 每根肌纖維僅由一個運動神經元支配。 肌肉中由單一運動神經元支配的〝實際肌纖維組〞稱為運動單位〔motor unit〕。 〝運動單位的大小〞會根據〝肌肉的性質〞而改變。
小運動單位是一種排列〔arrangement〕,其中單一運動神經元提供肌肉中的少量肌纖維。 小運動單位允許對肌肉進行非常精細的運動控制。 人類最好的例子是〝移動眼球的眼外肌〞的小運動單位。 每塊肌肉中有數千根肌纖維,但每六根左右的肌纖維由單一運動神經元提供,因為軸突分支在各自的神經肌肉接合(NMJ)形成突觸連接。 這允許眼球運動的精確控制,使雙眼能夠快速聚焦在同一物體上。 小運動單位也參與手指和拇指的許多精細運動,用於抓握、發短信等。
大運動單位是一種排列〔arrangement〕,其中單一運動神經元提供〝一個肌肉中的大量肌肉纖維〞。 大運動單位涉及簡單的或“粗略的〔gross〕”運動,例如有力地伸展膝關節。 最好的例子是大腿肌肉或背部肌肉的大運動單位,其中單一運動神經元將提供〝一個肌肉中的數千個肌肉纖維〞,因為它的軸突分裂成數千個分支。
許多骨骼肌內都有各種各樣的運動單位,這使得神經系統對肌肉有廣泛的控制能力。 肌肉中的小運動單位將具有較小、閾值較低的運動神經元,它們更容易興奮,首先向骨骼肌纖維放電,而骨骼肌纖維往往也是最小的。 這些〝較小運動單位的激活〞導致肌肉產生相對較小程度的收縮強度(張力)。 當需要更大的力量時,更大的運動單位以及更大、閾值更高的運動神經元就會被用來激活更大的肌肉纖維。 運動單位激活的增加會導致肌肉收縮增加,稱為募集〔recruitment〕。 隨著更多的運動單位被招募,肌肉收縮逐漸增強。 在某些肌肉中,最大的運動單位所產生的收縮力可能是肌肉中最小運動單位的 50 倍。 這樣可以使用肱二頭肌以最小的力量拾取羽毛,以及透過招募最大的運動單位由同一塊肌肉舉起重物。
必要時,可以同時招募肌肉中最大數量的運動單位,從而為該肌肉產生最大的收縮力,但由於維持收縮所需的能量,這種情況不能持續很長時間。 為了防止肌肉完全疲勞,運動單位通常不會全部同時活動,而是一些運動單位休息,而其他運動單位則活動,從而允許更長的肌肉收縮。 神經系統利用募集作為機制,以有效地利用骨骼肌。
肌節的長度-張力範圍
當骨骼肌纖維收縮時,肌球蛋白頭〔myosin heads〕附著在肌動蛋白〔actin〕上形成橫橋〔cross-bridges〕,隨後當肌球蛋白頭〔myosin heads〕拉動肌動蛋白〔actin〕時,細絲在粗絲上滑動,這導致肌節縮短,產生肌肉收縮的張力。 橫橋只能在細絲和粗絲已經重疊的地方形成,因此〝肌節的長度〞對〝肌節縮短時產生的力量〞有直接影響。 這稱為長度-張力關係〔length-tension relationship〕。
肌節產生最大張力的理想長度發生在其靜止長度的 80% 至 120%,其中 100% 是〝細絲的內側邊緣〞恰好位於〝粗絲的最內側肌球蛋白頭〞的狀態(圖 10.14)。 這個長度最大化了肌動蛋白結合位點〔actin-binding sites〕和肌球蛋白頭〔myosin heads〕的重疊。 如果肌節拉伸超過這個理想長度(超過 120%),粗絲和細絲就不會充分重疊,導致產生的張力較小。 如果肌節縮短超過 80%,重疊區域就會減少,伴隨著細絲伸出最後一個肌球蛋白頭,並且縮小了H區(H 區通常由肌球蛋白尾〔myosin tails〕組成)。 最終,細絲無處可去,張力也隨之減少。 如果肌肉被伸展到粗絲和細絲完全不重疊的程度,則無法形成橫橋,並且該肌節不會產生張力。 這種程度的伸展通常不會發生,因為輔助蛋白〔accessory proteins〕和結締組織會阻止過度伸展。
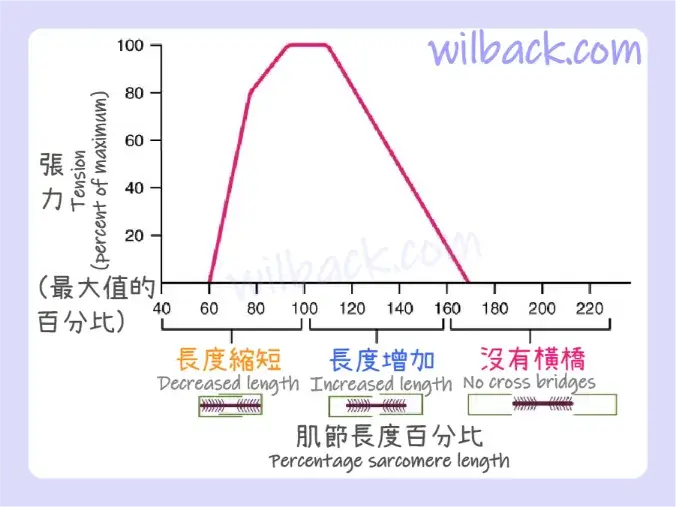
當粗絲和細絲重疊約 80% 至 120% 時,肌節會產生最大張力。
運動神經元刺激的頻率
〝來自運動神經元的單一動作電位〞將在其運動單位的肌肉纖維中產生單次收縮。 這種孤立的收縮稱為抽搐〔twitch〕。 抽搐可以持續幾毫秒或 100 毫秒,具體取決於肌肉類型。 單次抽搐產生的張力可以透過肌電圖來測量,肌電圖〔myogram〕是一種測量隨時間推移產生的張力大小的儀器(圖 10.15)。 每次抽搐都會經歷三個階段。 第一階段是潛伏期〔latent period〕,在此期間動作電位沿著肌膜傳播,並且Ca++離子從肌漿網 (SR)中釋放。 這是興奮和收縮耦合但收縮尚未發生的階段。 接下來發生收縮階段〔contraction phase〕。 肌漿中的 Ca++ 離子已與肌鈣蛋白〔troponin〕結合,原肌球蛋白〔tropomyosin〕已從肌動蛋白上的肌球蛋白結合位點〔myosin-binding sites〕轉移,橫橋已形成,且肌節正在主動縮短至峰值張力點。 最後一個階段是放鬆階段〔relaxation phase〕,此時張力會隨著收縮停止而減少。 Ca++ 離子從肌漿泵入肌漿網 (SR),橫橋循環停止,使肌纖維恢復到靜止狀態。
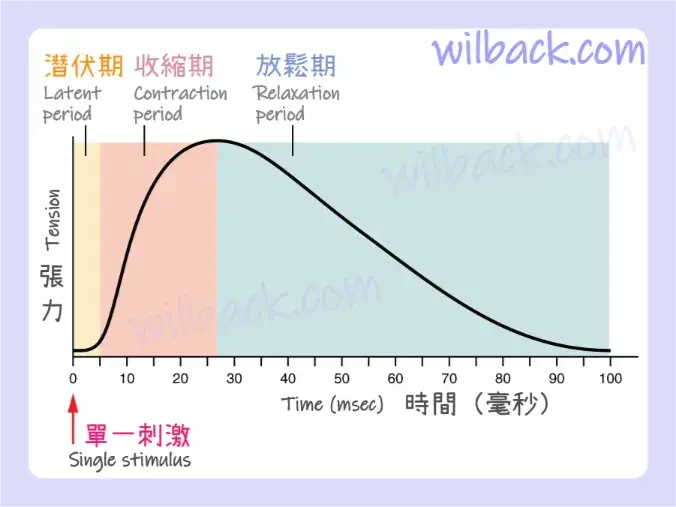
單一肌肉抽搐有一個潛伏期、張力增加時的收縮期和張力減少時的鬆弛期。 在潛伏期,動作電位沿著肌膜傳播。 在收縮階段,肌漿中的 Ca++ 離子與肌鈣蛋白〔troponin〕結合,原肌球蛋白〔tropomyosin〕從肌動蛋白結合位點〔actin-binding sites〕移動,肌動蛋白和肌球蛋白之間形成橫橋〔cross-bridges〕,並且肌節縮短。 在鬆弛階段,隨著 Ca++ 離子從肌漿中泵出並且橫橋循環停止,張力降低。
儘管人們可以經歷肌肉 “抽搐〔twitch〕”,但單次抽搐不會在活體中產生任何顯著的肌肉活動。 肌肉纖維需要一系列動作電位才能產生可做功的肌肉收縮。 正常的肌肉收縮更加持續,並且可以透過神經系統的輸入進行修改,以產生不同程度的力量; 這稱為分級肌肉反應〔graded muscle response〕。 〝來自運動神經元的動作電位(神經衝動)的頻率〞和〝傳遞動作電位的運動神經元的數量〞都會影響骨骼肌產生的張力。
〝運動神經元發出動作電位的速率〞會影響〝骨骼肌產生的張力〞。 如果在前一次抽搐仍在發生時刺激纖維,則第二次抽搐會更強。 這種反應稱為波加成〔wave summation〕,因為連續運動神經元訊號傳導的興奮-收縮耦聯〔excitation-contraction coupling〕效應被加成或相加(圖 10.16a)。 在分子層面上,加成的發生是因為第二次刺激觸發了更多 Ca++ 離子的釋放,這些離子可在〝肌肉仍因第一次刺激而收縮時〞激活額外的肌節。 加成導致運動單位的更大收縮。
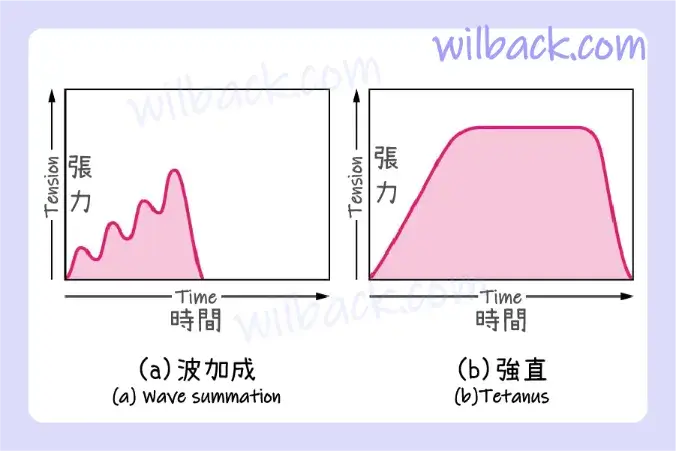
(a)連續運動神經元訊號傳導的興奮-收縮耦聯〔excitation-contraction coupling〕效應被加在一起,稱為波加成〔wave summation〕。 每個波的底部,即放鬆階段的結束,代表刺激點。 (b) 當刺激頻率太高以致鬆弛階段完全消失時,收縮變得連續; 這稱為強直〔tetanus〕。
如果運動神經元訊號傳導的頻率增加,〝運動單位中的總和〞和〝隨後的肌肉張力〞會繼續上升,直到達到尖峰點。 〝此尖峰點的張力〞大約比〝單次抽搐的張力〞大三到四倍,這種狀態稱為不完全強直〔incomplete tetanus〕。 在不完全強直期間,肌肉會經歷快速的收縮週期,每個週期都有短暫的放鬆階段。 如果刺激頻率太高以至於鬆弛階段完全消失,則收縮會在一個稱為完全強直〔complete tetanus〕的過程中變得連續(圖 10.16b)。
在強直期間,〝肌漿中 Ca++ 離子的濃度〞允許幾乎所有肌節形成橫橋並縮短,因此收縮可以不間斷地持續(直到肌肉疲勞並且不再產生張力)。
階梯現象
當骨骼肌休眠很長一段時間然後激活收縮時,在所有其他條件相同的情況下,最初的收縮產生的力量大約是後來收縮的一半。 肌肉張力逐漸增加,有點看起來像一組樓梯。 這種張力增加被稱為階梯現象〔treppe〕,這是一種肌肉收縮變得更有效的情況。 它也被稱為 “階梯效應〔staircase effect〕”(圖 10.17)。
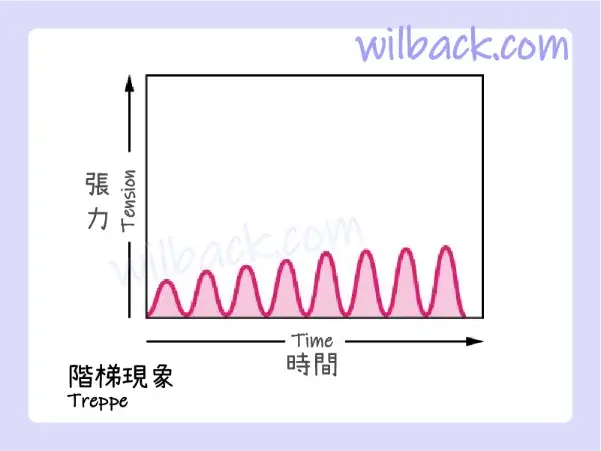
當肌肉張力逐漸增加(看起來像一組樓梯)時,稱為階梯現象〔treppe〕。 每個波浪的底部代表刺激點。
據信,階梯現象〔treppe〕是由於來自運動神經元的穩定訊號流導致〝肌漿中 Ca++ 濃度較高〞所致。 只有足夠的 ATP 才能維持它。
肌肉張力
骨骼肌很少完全放鬆或鬆弛。 即使肌肉不產生運動,它也會少量收縮以維持其收縮蛋白,並且產生肌張力〔muscle tone〕。 肌張力產生的張力使肌肉能夠持續穩定關節並保持姿勢。
肌肉張力是透過神經系統和骨骼肌之間複雜的相互作用來實現的,這導致一次激活幾個運動單位(很可能以週期性方式激活)。 透過這種方式,肌肉永遠不會完全疲勞,因為有些運動單位可以恢復,而有些則是活躍。
缺乏〝導致肌張力的低水平收縮〞被稱為張力減退〔hypotonia〕,可能是由於中樞神經系統 (CNS) 部分(例如小腦)受損或骨骼肌神經支配喪失(例如小兒麻痺症)所致。 低張力肌肉外觀鬆弛,並表現出功能障礙,例如弱反射〔weak reflexes〕。 相反,過度的肌肉張力被稱為張力亢進〔hypertonia〕,並伴隨反射亢進〔hyperreflexia〕(過度反射反應),通常是中樞神經系統上運動神經元受損的結果。 肌張力亢進可表現為肌肉僵硬(如帕金森氏症)或痙攣(即肌張力的階段性變化),肢體會從被動伸展中 “彈回〔snap〕”(如某些中風)。
更新紀錄
2024/03/17 發佈本文
評論