

 夜黎
夜黎本文章內容基於萊斯大學 OpenStax 的 Anatomy and Physiology 2e,由夜黎重新編輯。(根據本書前言中的創用 CC BY 4.0 聲明)
原文傳送門:<10> Muscle Tissue — 10.3 Muscle Fiber Contraction and Relaxation
索引傳送門:《解剖學和生理學2e》索引頁面
學習本節後,你將能夠:
- 描述肌肉收縮的組成部分
- 解釋肌肉如何收縮、放鬆
- 描述肌肉收縮的肌絲滑動〔sliding filament〕模型
前言
導致〝單一肌纖維收縮的事件序列〞始於一個信號——神經遞質乙醯膽鹼(ACh)——來自支配該纖維的運動神經元。 當帶正電的鈉離子 (Na+) 進入時,纖維的局部膜將去極化,觸發一個動作電位〔action potential〕,該動作電位會擴散到膜的其餘部分,包括 T 小管〔T-tubules〕,使之去極化。 這會觸發肌漿網 (SR) 中儲存的鈣離子 (Ca++) 的釋放。 然後 Ca++ 開始收縮,由三磷酸腺苷(ATP) 維持(圖 10.8)。 只要 Ca++ 離子保留在肌漿中與肌鈣蛋白結合,從而使肌動蛋白結合位點保持 “未屏蔽〔unshielded〕”,並且只要 ATP 可用於驅動〝橫橋循環〔cross-bridge cycling〕〞和〝肌球蛋白對肌動蛋白絲線〔actin strands〕的牽引〞,肌纖維將繼續縮短至解剖學極限。
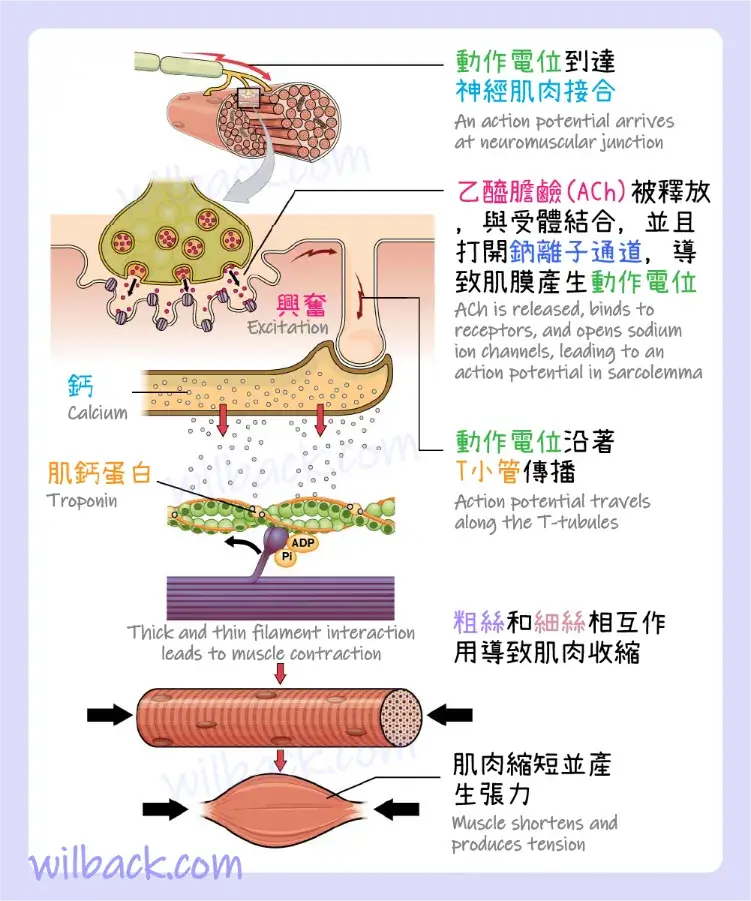
肌動蛋白和肌球蛋白頭之間形成橫橋,觸發收縮。 只要 Ca++ 離子保留在肌漿中與肌鈣蛋白結合,以及只要有 ATP 可用,肌纖維就會繼續縮短。
當來自運動神經元的訊號結束時,肌肉收縮通常會停止,這會使肌纖維膜和 T 小管重新極化,並關閉肌漿網 (SR)中的電壓門控鈣通道〔voltage-gated calcium channels〕。 然後 Ca++ 離子被泵回肌漿網 (SR),導致原肌球蛋白〔tropomyosin〕重新屏蔽(或重新覆蓋)肌動蛋白絲線〔actin strands〕上的結合位點。 當三磷酸腺苷(ATP)耗盡並變得疲勞時,肌肉也會停止收縮(圖 10.9)。
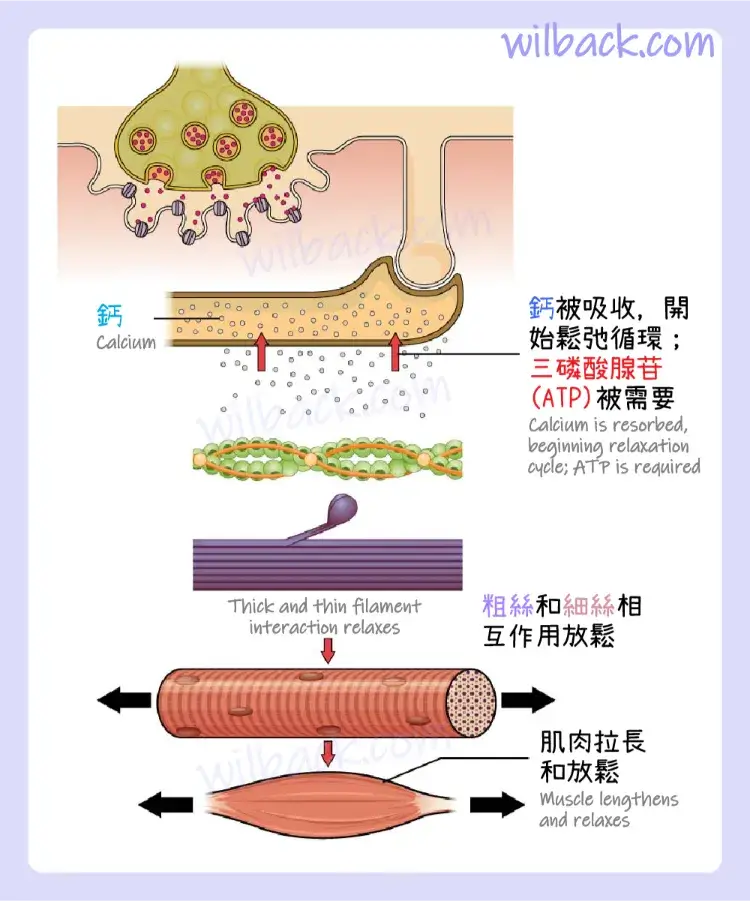
Ca++ 離子被泵回 SR,導致原肌球蛋白重新屏蔽肌動蛋白絲線上的結合位點。 當 ATP 耗盡並變得疲勞時,肌肉也可能會停止收縮。
肌纖維縮短的分子事件發生在纖維的肌節內(見圖 10.10)。 當〝肌原纖維〔myofibrils〕內線性排列的肌節〔sarcomeres〕〞隨著肌球蛋白頭〔myosin heads〕拉動肌動蛋白絲〔actin filaments〕而縮短時,橫紋肌纖維就會收縮。
粗絲和細絲重疊的區域具有緻密的外觀,因為絲之間的空間很小。 〝細絲和粗絲重疊的區域〞對於肌肉收縮非常重要,因為它是絲運動開始的部位。 細絲的末端由 Z 盤〔Z-discs〕錨定,不會完全延伸到〝僅包含粗絲的中心區域〞,粗絲的基部錨定在稱為 M 線〔M-line〕的點處。 肌原纖維〔myofibril〕由許多沿其長度延伸的肌節〔sarcomeres〕組成。 因此,肌原纖維和肌肉細胞會隨著肌節的收縮而收縮。
收縮的肌絲滑動模型
當運動神經元發出訊號時,骨骼肌纖維會隨著細絲被拉動而收縮,然後滑動纖維肌節內的粗絲。 這個過程稱為肌肉收縮的肌絲滑動模型〔sliding filament model〕(圖10.10)。 只有當〝肌動蛋白絲〔actin filaments〕上的肌球蛋白結合位點〔myosin-binding sites〕〞透過一系列步驟(始於 Ca++ 進入肌漿)暴露時,才會發生滑動。
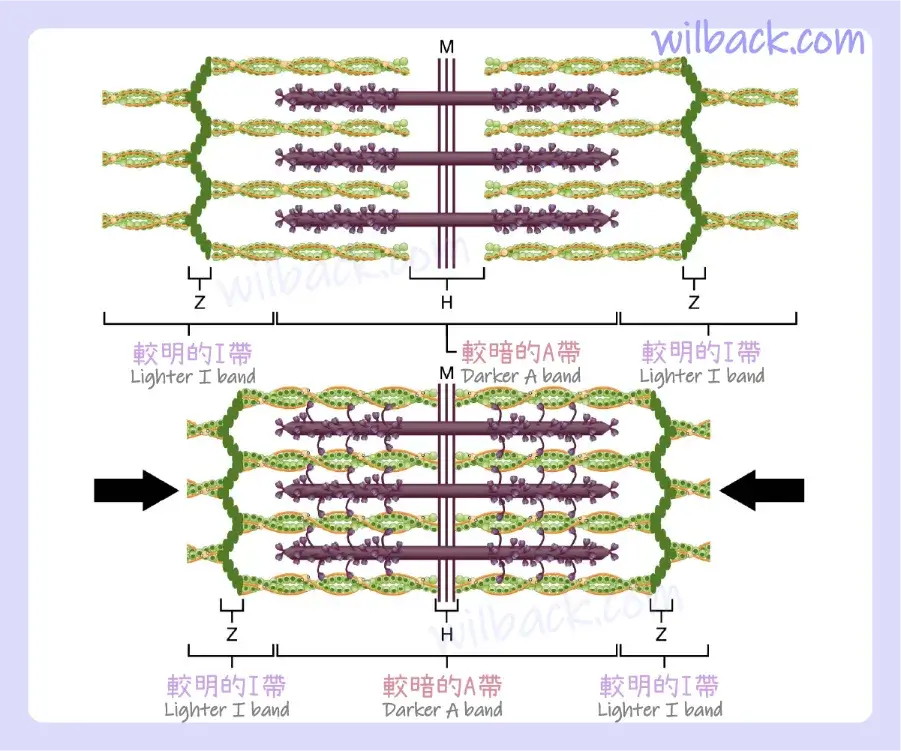
當肌節收縮時,Z 線靠得更近,並且I 帶變小。 A 帶保持相同的寬度。 完全收縮時,細絲和粗絲完全重疊。
原肌球蛋白〔tropomyosin〕是一種蛋白質,纏繞在肌動蛋白絲〔actin filament〕的鏈上,並覆蓋肌球蛋白結合位點〔myosin-binding sites〕,以防止肌動蛋白〔actin〕與肌球蛋白〔myosin〕結合。 原肌球蛋白〔tropomyosin〕與肌鈣蛋白結合〔troponin〕形成肌鈣蛋白-原肌球蛋白複合物。 肌鈣蛋白-原肌球蛋白複合物〔troponin-tropomyosin complex〕可防止肌球蛋白〔myosin〕 “頭部〔heads〕” 與肌動蛋白微絲〔actin microfilaments〕上的活性位點結合。 肌鈣蛋白也具有 Ca++ 離子的結合位點。
為了啟動肌肉收縮,原肌球蛋白〔tropomyosin〕必須暴露肌動蛋白絲〔actin filament〕上的肌球蛋白結合位點〔myosin-binding site〕,以允許肌動蛋白和肌球蛋白微絲〔actin and myosin microfilaments〕之間形成橫橋〔cross-bridge〕。 收縮過程的第一步是〝Ca++ 與肌鈣蛋白結合〞,以便原肌球蛋白〔tropomyosin〕可以滑離肌動蛋白絲線〔actin strands〕上的結合位點。 這使得肌球蛋白頭〔myosin heads〕能夠結合到這些暴露的結合位點並形成橫橋。 然後細絲被肌球蛋白頭〔myosin heads〕拉動,滑過粗絲滑向肌節的中心。 但是,每個頭部只能拉動非常短的距離,就會達到其極限,它必須 “重新翹起〔re-cocked〕”才能再次拉動,這一步驟需要三磷酸腺苷(ATP)。
三磷酸腺苷(ATP)和肌肉收縮
為了使細絲在肌肉收縮期間繼續滑過粗絲,肌球蛋白頭〔myosin heads〕必須在結合位點拉動肌動蛋白〔actin〕,然後分離、重新翹起、附著到更多結合位點、拉動、分離、重新翹起等等。 這種重複的運動稱為橫橋循環〔cross-bridge cycle〕。 肌球蛋白頭〔myosin heads〕的這種移動類似於個人划船時的槳:槳的槳葉(肌球蛋白頭)拉動,從水中提起(分離),重新定位(重新翹起),然後再次浸入水中拉動(圖10.11 )。 每個循環都需要能量,而肌節中的肌球蛋白頭〔myosin heads〕重複拉動細絲的動作也需要能量,而能量由三磷酸腺苷(ATP)提供。
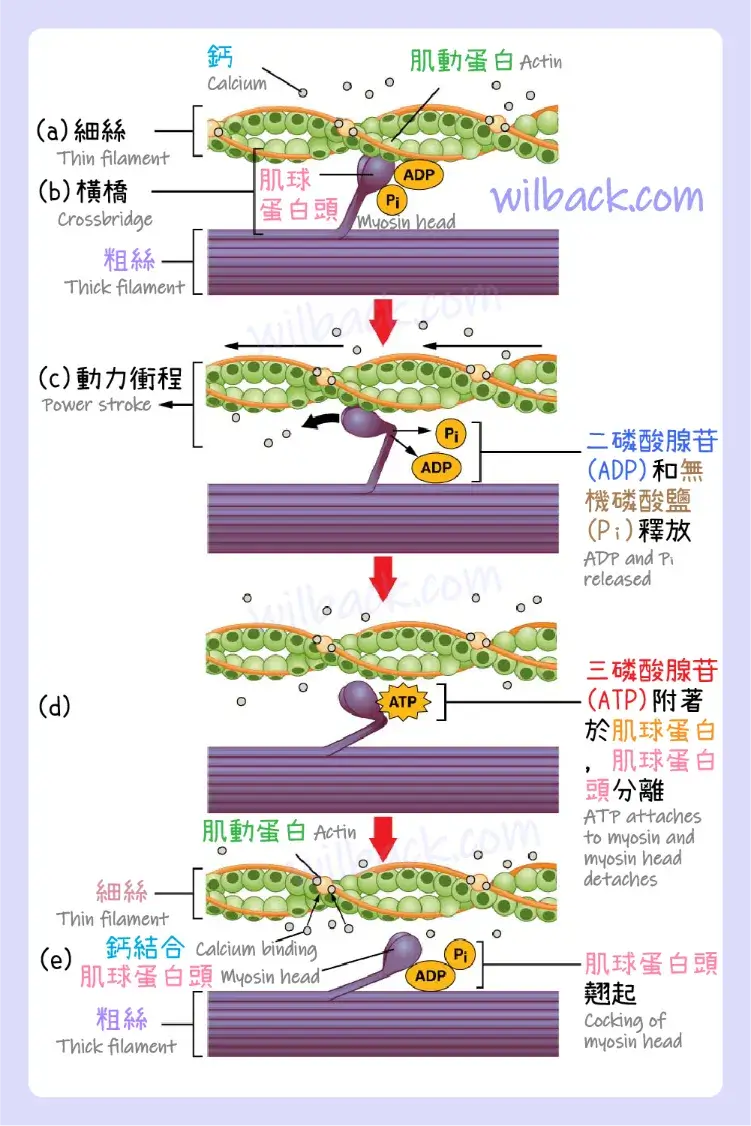
(a) 當鈣與肌鈣蛋白結合時,肌動蛋白上的活性位點暴露。 (b) 肌球蛋白頭被肌動蛋白吸引,並且肌球蛋白在其肌動蛋白結合位點結合肌動蛋白,形成橫橋。 (c) 在動力衝程期間,前一個收縮週期所產生的磷酸鹽會被釋放。 這導致肌球蛋白頭部向肌節中心旋轉,然後釋放附著的 ADP 和磷酸基團。 (d) 一個新的 ATP 分子附著在肌球蛋白頭,導致橫橋分離。 (e) 肌球蛋白頭將 ATP 水解為 ADP 和磷酸鹽,使肌球蛋白回到翹起位置。
當肌球蛋白頭〔myosin head〕附著於肌動蛋白〔actin〕,而二磷酸腺苷(ADP)和無機磷酸鹽(Pi)仍與肌球蛋白結合時,就會形成橫橋〔cross-bridge〕(圖 10.11a、b)。 然後無機磷酸鹽(Pi)被釋放,導致肌球蛋白對肌動蛋白形成更強的附著力,之後肌球蛋白頭向 M 線移動,同時拉動肌動蛋白。 當肌動蛋白被拉動時,絲向 M 線移動約 10 nm。 這種運動稱為動力衝程〔power stroke〕,因為細絲的運動發生在這一步驟(圖 10.11c)。 在沒有三磷酸腺苷(ATP)的情況下,肌球蛋白頭不會與肌動蛋白分離。
〝肌球蛋白頭的一部分〞附著在〝肌動蛋白上的結合位點〞,但該頭部還有另一個三磷酸腺苷(ATP)結合位點。 三磷酸腺苷(ATP)結合導致肌球蛋白頭與肌動蛋白分離(圖 10.11d)。 發生這種情況後,三磷酸腺苷(ATP)會透過肌球蛋白的內在三磷酸腺苷酶〔ATPase〕活性轉化為二磷酸腺苷(ADP)和無機磷酸鹽(Pi)。 〝三磷酸腺苷(ATP)水解過程中釋放的能量〞將肌球蛋白頭的角度改為翹起位置(圖 10.11e)。 肌球蛋白頭現在處於可以進一步移動的位置。
當肌球蛋白頭翹起時,肌球蛋白處於高能構型。 當肌球蛋白頭移動通過動力衝程時,該能量被消耗,並且在動力衝程結束時,肌球蛋白頭處於低能量位置。 動力衝程後,二磷酸腺苷(ADP)被釋放; 然而,形成的橫橋仍然存在,並且肌動蛋白和肌球蛋白結合在一起。 只要三磷酸腺苷(ATP)可用,它就很容易附著在肌球蛋白上,橫橋循環就可以重複,肌肉收縮就可以持續。
請注意,每根粗絲由大約 300 個肌球蛋白分子組成,具有多個肌球蛋白頭,並且許多橫橋在肌肉收縮過程中不斷形成和斷裂。 將其乘以〝一根肌原纖維中的所有肌節〞、〝一根肌纖維中的所有肌原纖維〞以及〝一根骨骼肌中的所有肌纖維〞,你就可以理解為什麼需要如此多的能量—三磷酸腺苷(ATP)—來保持骨骼肌的工作。 事實上,正是三磷酸腺苷(ATP)的損失導致了人類死後不久就會出現屍僵。 由於無法進一步產生三磷酸腺苷(ATP),因此肌球蛋白頭無法從肌動蛋白結合位點上分離出三磷酸腺苷(ATP),因此橫橋保持在原位,導致骨骼肌僵化。
三磷酸腺苷(ATP)的來源
三磷酸腺苷(ATP)為肌肉收縮提供能量。 除了在橫橋循環中的直接作用外,三磷酸腺苷(ATP)還為肌漿網(SR)中的〝主動運輸 Ca++ 幫浦〔active-transport Ca++ pumps〕〞提供能量。 如果沒有足夠的三磷酸腺苷(ATP)量,肌肉就不會收縮。 肌肉中儲存的三磷酸腺苷(ATP)量非常低,僅足以為幾秒鐘的收縮提供動力。 因此,當三磷酸腺苷(ATP)被分解時,它必須快速再生和替換,以實現持續收縮。 肌肉細胞中三磷酸腺苷(ATP)再生的機制有三種:磷酸肌酸代謝〔phosphate metabolism〕、無氧糖解反應〔anaerobic glycolysis〕和有氧呼吸〔aerobic respiration〕。
肌酸磷酸〔creatine phosphate〕是一種可以在其磷酸鍵中儲存能量的分子。 在休息的肌肉中,過量的三磷酸腺苷(ATP)將其能量轉化為肌酸〔creatine〕,產生三磷酸腺苷(ATP)和肌酸磷酸〔creatine phosphate〕。 這充當能量儲備,可用於快速產生更多三磷酸腺苷(ATP)。 當肌肉開始收縮並需要能量時,肌酸磷酸〔creatine phosphate〕會將其磷酸鹽轉移回二磷酸腺苷(ADP),形成三磷酸腺苷(ATP)和肌酸〔creatine〕。 此反應由肌酸激酶〔creatine kinase〕催化,發生得非常快; 因此,肌酸磷酸〔creatine phosphate〕衍生的三磷酸腺苷(ATP)為肌肉收縮的前幾秒提供動力。 然而,肌酸磷酸〔creatine phosphate〕只能提供約 15 秒的能量,此時必須使用另一種能量資源(圖 10.12)。
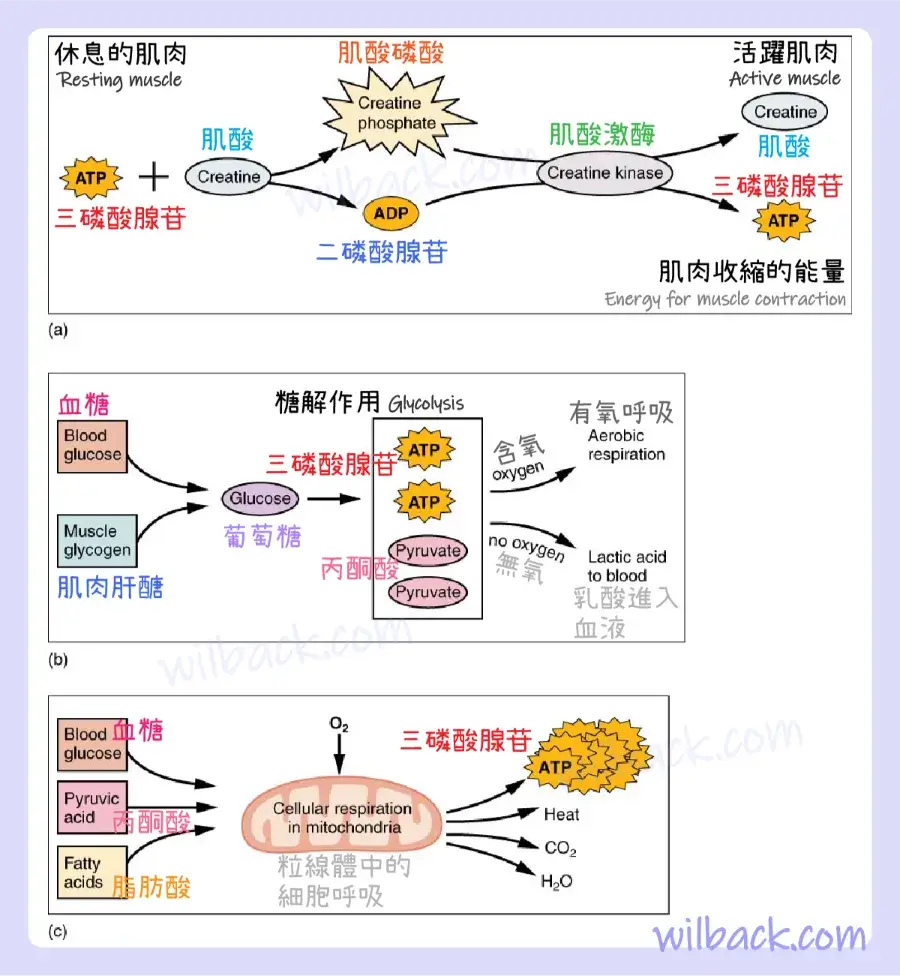
(a) 有些 ATP 儲存在靜止的肌肉中。 當收縮開始時,它會在幾秒鐘內耗盡。 約 15 秒內,磷酸肌酸會產生更多 ATP。 (b) 每個葡萄糖分子產生兩個 ATP 和兩個丙酮酸〔pyruvic acid〕分子,可用於有氧呼吸或轉化為乳酸〔lactic acid〕。 如果沒有氧氣,丙酮酸會轉化為乳酸,這可能會導致肌肉疲勞。 這種情況發生在劇烈運動期間,當需要大量能量但氧氣無法充分輸送到肌肉時。 (c) 有氧呼吸是指葡萄糖在氧氣 (O2) 存在下分解,產生二氧化碳、水和 ATP。 休息或適度活動的肌肉所需的 ATP 大約 95% 由發生在粒線體中的有氧呼吸提供。
隨著肌酸磷酸〔creatine phosphate〕產生的三磷酸腺苷(ATP)耗盡,肌肉轉向糖解作用〔glycolysis〕作為三磷酸腺苷(ATP)來源。 糖解作用〔glycolysis〕是一種無氧(不依賴氧氣)過程,分解葡萄糖(糖)以產生三磷酸腺苷(ATP); 然而,糖解作用不能像肌酸磷酸〔creatine phosphate〕那樣快速產生三磷酸腺苷(ATP)。 因此,轉向糖解作用會導致肌肉的三磷酸腺苷(ATP)利用率降低。 糖解作用〔glycolysis〕中使用的糖可以由〝血糖〞或〝經由代謝儲存在肌肉中的肝醣〔glycogen〕〞提供。 一個葡萄糖分子的分解產生兩個三磷酸腺苷(ATP)和兩個丙酮酸〔pyruvic acid〕分子,可用於有氧呼吸或在氧氣水平較低時轉化為乳酸〔lactic acid〕(圖 10.12b)。
如果有氧氣可用,丙酮酸〔pyruvic acid〕用於有氧呼吸。 然而,如果沒有氧氣可用,丙酮酸〔pyruvic acid〕就會轉化為乳酸〔lactic acid〕,這可能會導致肌肉疲勞。 這種轉化允許從〝菸鹼醯胺腺嘌呤二核苷酸(NADH)〞中回收〝菸醯胺腺嘌呤二核酸(NAD+)〞酶,這是糖解作用繼續進行所必需的。 這種情況發生在劇烈運動期間,當需要大量能量但氧氣無法充分輸送到肌肉時。 糖解作用本身不能持續很長時間(大約 1 分鐘的肌肉活動),但它有助於促進短時間的高強度輸出。 這是因為糖解作用無法非常有效地利用葡萄糖,每個葡萄糖分子會淨增加兩個三磷酸腺苷(ATP),並最終產生乳酸〔lactic acid〕,而乳酸的累積可能會導致肌肉疲勞。
有氧呼吸〔aerobic respiration〕是葡萄糖或其他營養物質在氧氣 (O2) 的存在下分解,產生二氧化碳、水和三磷酸腺苷(ATP)。 〝休息或適度活動的肌肉所需的三磷酸腺苷(ATP)〞大約 95% 由發生在粒線體中的有氧呼吸提供。 有氧呼吸的輸入包括血液中循環的葡萄糖〔glucose〕、丙酮酸〔pyruvic acid〕和脂肪酸〔fatty acids〕。 有氧呼吸比無氧糖解作用效率高得多,每個葡萄糖分子產生約 36 個三磷酸腺苷(ATP),而糖解則產生 4 個三磷酸腺苷(ATP)。 然而,如果骨骼肌沒有穩定的氧氣供應,有氧呼吸就無法持續,而且速度要慢得多(圖 10.12c)。 為了補償,肌肉在稱為肌紅蛋白〔myoglobin〕的蛋白質中儲存少量過量的氧氣,使肌肉收縮更有效並減少疲勞。 有氧訓練還可以提高循環系統的效率,從而可以更長時間地向肌肉提供氧氣。
當肌肉無法再響應神經系統的訊號而收縮時,就會發生肌肉疲勞。 儘管某些因素與疲勞期間發生的肌肉收縮減少有關,但肌肉疲勞的確切原因尚不完全清楚。 正常肌肉收縮需要三磷酸腺苷(ATP),隨著三磷酸腺苷(ATP)儲備的減少,肌肉功能可能會下降。 這可能更多是短暫、強烈的肌肉輸出的一個因素,而不是持續、較低強度的努力的因素。 乳酸〔lactic acid〕的累積可能會降低細胞內的 pH 值,影響酵素和蛋白質的活性。 膜去極化〔membrane depolarization〕導致的 Na+ 和 K+ 水平不平衡可能會擾亂 Ca++ 流出肌漿網(SR)。 長時間持續運動可能會損害肌漿網(SR)和肌膜〔sarcolemma〕,導致 Ca++ 調節受損。
劇烈的肌肉活動會導致氧債〔oxygen debt〕,即補償肌肉收縮期間無氧時產生的三磷酸腺苷(ATP) 所需的氧氣量。 需要氧氣來恢復三磷酸腺苷(ATP)和肌酸磷酸〔creatine phosphate〕水平,將乳酸〔lactic acid〕轉化為丙酮酸〔pyruvic acid〕,並在肝臟中將乳酸轉化為葡萄糖〔glucose〕或肝醣〔glycogen〕。 運動期間使用的其他系統也需要氧氣,所有這些組合過程都會導致運動後呼吸頻率增加。 在滿足氧債〔oxygen debt〕之前,即使運動停止後,氧氣攝取量也會增加。
骨骼肌的鬆弛
放鬆骨骼肌纖維,最終放鬆骨骼肌,從運動神經元開始,運動神經元停止向神經肌肉接合(NMJ)突觸釋放化學訊號乙醯膽鹼(ACh)。 肌纖維將重新極化,從而關閉肌漿網(SR)中釋放 Ca++ 的門閥。 三磷酸腺苷(ATP)驅動的幫浦會將 Ca++ 從肌漿中移回肌漿網(SR)中。 這導致細絲上肌動蛋白結合位點的 “重新屏蔽〔reshielding〕”。 如果沒有能力在細絲和粗絲之間形成橫橋,肌纖維就會失去張力放鬆。
肌肉力量
給定肌肉中骨骼肌纖維的數量是由基因決定的,並且不會改變。 肌肉力量與〝每根纖維內的肌原纖維和肌節的數量〞有直接關係。 作用於肌肉的激素和壓力(以及人工合成代謝類固醇)等因素可以增加肌纖維內肌節〔sarcomeres〕和肌原纖維〔myofibrils〕的產生,這種變化稱為肥大〔hypertrophy〕,導致骨骼肌的質量和體積增加。 同樣,骨骼肌使用的減少會導致萎縮,即肌節和肌原纖維的數量消失(但肌纖維的數量不會消失)。 拆除石膏後,打著石膏的肢體通常會出現肌肉萎縮,而某些疾病,例如小兒麻痺,也會出現肌肉萎縮。
肌肉系統
裘馨氏肌肉失養症 (DMD) 〔Duchenne muscular dystrophy〕是一種骨骼肌進行性衰弱的疾病。 它是統稱為 “肌肉失養症〔muscular dystrophy〕” 的幾種疾病之一。 DMD 是由於肌失養蛋白〔dystrophin〕引起的,這種蛋白有助於肌原纖維的細絲〔thin filaments〕與肌膜〔sarcolemma〕結合。 如果沒有足夠的肌失養蛋白〔dystrophin〕,肌肉收縮會導致肌膜撕裂,引起 Ca++ 流入,導致細胞損傷和肌纖維降解。 隨著時間的推移,隨著肌肉損傷的累積,肌肉質量會流失,並且會出現更大的功能障礙。
DMD 是一種由 X 染色體異常引起的遺傳性疾病。 它主要影響男性,通常在兒童早期被診斷出來。 DMD 通常首先表現為平衡和運動困難,然後發展為無法行走。 它在身體中繼續從下肢向上發展到上半身,影響負責呼吸和循環的肌肉。 它最終會導致呼吸衰竭而死亡,而患者通常活不過 20 多歲。
由於 DMD 是由編碼肌失養蛋白〔dystrophin〕的基因突變引起的,因此人們認為將健康的肌母細胞〔myoblasts〕引入患者體內可能是一種有效的治療方法。 肌母細胞〔myoblasts〕是負責肌肉發育的胚胎細胞,理想情況下,它們攜帶健康的基因,可以產生正常肌肉收縮所需的肌失養蛋白〔dystrophin〕。 這種方法在人類身上基本上不成功。 最近的一種方法涉及嘗試促進肌肉產生〝肌失養蛋白相關蛋白〔utrophin〕〞,這是一種類似於肌失養蛋白〔dystrophin〕的蛋白質,可能能夠承擔肌失養蛋白的作用並防止細胞損傷的發生。
更新紀錄
2024/03/01 發佈本文
評論